Magnetotactic bacteria (MTB) are a diverse group of prokaryotes that have a singular ability to align with geomagnetic field lines. This ability is due to special organelles called magnetosomes. Magnetosomes are composed of single-magnetic-domain nanocrystals of magnetite [Fe(II)Fe(III)2O4] or greigite [Fe(II)Fe(III)2S4] embedded in biological membrane.
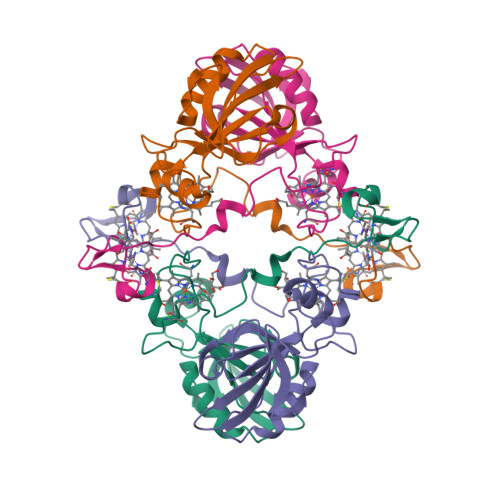
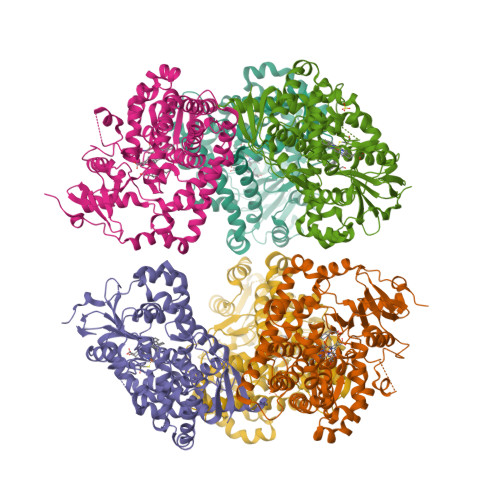
“Magnetochrome” is a name proposed in 2012 by Marina Siponen and co-authors for a cytochrome domain conserved within all known MTB and not found in any other species to date [1]. Recently, the crystal structure of the magnetosome-associated protein MamP has been solved at 1.8 Å resolution [2—4]. The minimal unit of MamP is a dimer. Each monomer consists of a PDZ domain fused to two magnetochrome domains. It was also shown in an in vitro mineralisation experiment that MamP functions as an iron oxidase mediating the iron(III) ferrihydrite production from iron(II) [2]:
4Fe2+ + 7H2O → 2Fe2O3·H2O + 12H+ + 4e–
Siponen, M.I., Adryanczyk, G., Ginet, N., Arnoux, P. and Pignol, D. (2012) Magnetochrome: a c-type cytochrome domain specific to magnetotatic bacteria. Biochemical Society Transactions40, 1319—1323.
Siponen, M.I., Legrand, P., Widdrat, M., Jones, S.R., Zhang, W.-J., Chang, M.C.Y., Faivre, D., Arnoux, P. and Pignol, D. (2013) Structural insight into magnetochrome-mediated magnetite biomineralization. Nature502, 681—684.
A unicellular green alga Ostreococcus tauri is the smallest (less than 1 μm in diameter) free-living eukaryote yet described. Viruses that can infect high-light and low-light adapted strains of O. tauri have been isolated and their genomes sequenced. Interestingly, low-light-strain infecting virus (OtV-2) differ from the high-light-strain infecting viruses by encoding a potential cytochrome b5 [1]. This protein was cloned, biochemically characterised and its three-dimensional structure resolved [2, 3].
The absorption spectra of oxidised and reduced recombinant OtV-2 haemoprotein are almost identical to those of purified human cytochrome b5.
Absorbance spectra of purified recombinant human cytochrome b5 and OtV-2_201.
It was also shown that the protein can substitute for yeast cytochrome b5 in the CYP51-mediated sterol 14α-demethylation. Structurally, the viral protein is similar to other known cytochromes b5 but lacks a hydrophobic C-terminal anchor. Thus, the first virally encoded cytochrome b5 is also the first cytosolic cytochrome b5 characterised. However, the physiological role of viral cytochrome b5 remains unknown.
Known scientifically as hematite, ochre is a reddish iron-containing rock that was used as a coloring substance made by grinding the stone into a powder that when mixed with fat or water formed a paste. Homo erectus had already begun to work with ochre, thus securing it as a key, nonfunctional element in human life.
Ochre was a great universal of all First Societies. The Blackfeet of the American Plains referred to it as nitsisaan or “real paint” and profusely daubed it on their ceremonial garments. Its color was thought to represent the sun and the energy that permeates all things, making a person rubbed with it appear holy and powerful. Its redness and brilliance signaled supernatural potency overlapping with a range of cosmological concepts revolving around rain, fertility, hunting, and death. The nineteen-century painter George Catlin painted the Sioux worshipping at a red boulder in the open grasslands of the Great Plains.
George Catlin, Sioux Worshiping at the Red Boulders, 1837—1839
Ceremonial uses of ochre still exist today, such as among the Maasai in Kenya during certain initiation rituals, by Amazon tribes and by Aboriginal people in Australia.
The !Kung, who live in the Kalahari desert of Botswana and who are among the oldest of the surviving First Society people in the world, use the pigment in rituals dealing with a woman’s first menstruation. A female initiate, on emergence from seclusion, would present the women of her kin group with lumps of ochre for decorating their faces and cloaks and also for adorning the young men to protect them when out hunting.
This volume, edited by Lucia Banci, is probably the first real book on metallomics. The table of contents looks very promising, and judging from those bits that I am able to access, I’d love to say that is is a great book... But honestly I can’t. The days when I could persuade the library to purchase (for me) a book, however expensive, are long gone. At the Springer website, eBook is priced at €142.79 and hardcover costs €181.85. You can buy them slightly cheaper from Amazon ($175.82 and $227.05, respectively.)
The book is dedicated to Ivano Bertini, who sadly passed away last year. I was lucky enough to meet the man himself on a few occasions. Ivano was a formidable scientist and one of the most colourful figures of bioinorganic chemistry and structural biology.
Metallomics and the Cell Metal Ions in Life Sciences, vol. 12
Lucia Banci, Editor
Banci, L. and Bertini, I. Metallomics and the cell: some definitions and general comments, pp. 1—13.
Penner-Hahn, J.E. Technologies for detecting metals in single cells, pp. 15—40.
Clausen, M.J.V. and Poulsen, H. Sodium/potassium homeostasis in the cell, pp. 41—67.
Romani, A.M.P. Magnesium homeostasis in mammalian cells, pp. 69—118.
Brini, M., Calì, T., Ottolini, D. and Carafoli, E. Intracellular calcium homeostasis and signaling, pp. 119—168.
Roth, J., Ponzoni, S. and Aschner, M. Manganese homeostasis and transport, pp. 169—201.
Andrews, S., Norton, I., Salunkhe, A.S., Goodluck, H., Aly, W.S.M., Mourad-Agha, H. and Cornelis, P.
Control of iron metabolism in bacteria, pp. 203—239.
Dlouhy, A.C. and Outten, C.E. The iron metallome in eukaryotic organisms, pp. 241—278.
Benson, D.R. and Rivera, M. Heme uptake and metabolism in bacteria, pp. 279—332.
Cracan, V. and Banerjee, R. Cobalt and corrinoid transport and biochemistry, pp. 333—374.
Sydor, A.M. and Zamble, D.B. Nickel metallomics: general themes guiding nickel homeostasis, pp. 375—416.
Rensing, C. and McDevitt, S.F. The copper metallome in prokaryotic cells, 417—450.
Vest, K.E., Hashemi, H.F. and Cobine, P.A. The copper metallome in eukaryotic cells, pp. 451—478.
Maret, W. Zinc and the zinc proteome, pp. 479—501.
Mendel, R.R. Metabolism of molybdenum, pp. 503—528.
Gladyshev, V.N. and Zhang, Y. Comparative genomics analysis of the metallomes, pp. 529—580.
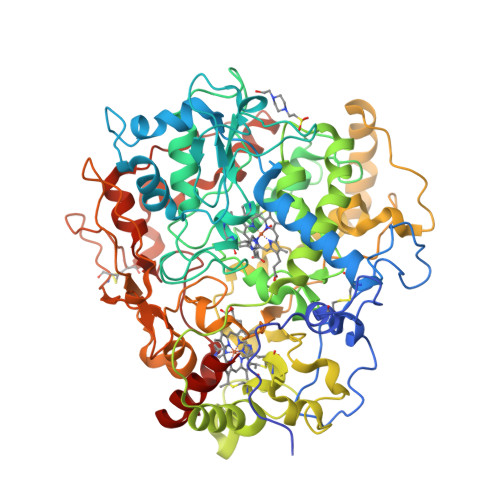
To date, two types of enzymes that are responsible for primary attack of polyisoprene in rubber-degrading microorganisms have been identified [1]. One is the latex clearing protein (Lcp), first isolated from Streptomyces sp., which does not have any metal ions or cofactors [2]. The other is the rubber oxygenase RoxA of Xanthomonas sp., a dihaem c-type cytochrome that cleaves cis-1,4-polyisoprene, the main constituent of natural rubber, to 12-oxo-4,8-dimethyltrideca-4,8-diene-1-al [3, 4]. The crystal structure of RoxA, solved at 1.8 Å resolution, was released today [5].
Rose, K., Tenberge, K.B. and Steinbüchel, A. (2005) Identification and characterization of genes from Streptomyces sp. strain K30 responsible for clear zone formation on natural rubber latex and poly(cis-1,4-isoprene) rubber degradation. Biomacromolecules6, 180—188.
Braaz, R., Fischer, P. and Jendrossek, D. (2004) Novel type of heme-dependent oxygenase catalyzes oxidative cleavage of rubber (poly-cis-1,4-isoprene). Applied and Environmental Microbiology70, 7388—7395.
Schmitt, G., Seiffert, G., Kroneck, P.M.H., Braaz, R. and Jendrossek, D. (2010) Spectroscopic properties of rubber oxygenase RoxA from Xanthomonas sp., a new type of dihaem dioxygenase. Microbiology156, 2537—2548.
A novel cofactor is not something that is discovered every day, or even every year. So we are lucky this year. The crystal structure of a putative methyltransferase CmoA from Escherichia coli reveals the presence of [(3S)-3-amino-3-carboxypropyl]{[(2S,3S,4R,5R)-5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl}(carboxymethyl)sulfanium, aka S-adenosyl-S-carboxymethyl-L-homocysteine, aka SCM-SAH [1—3]. Moreover, it was suggested that “a number of enzymes that have previously been annotated as SAM-dependent are in fact SCM-SAH-dependent” [1].
Byrne, R.T., Whelan, F., Aller, P., Bird, L.E., Dowle, A., Lobley, C.M., Reddivari, Y., Nettleship, J.E., Owens, R.J., Antson, A.A. and Waterman, D.G. (2013) S-Adenosyl-S-carboxymethyl-L-homocysteine: a novel cofactor found in the putative tRNA-modifying enzyme CmoA. Acta Crystallographica D69, 1090—1098.
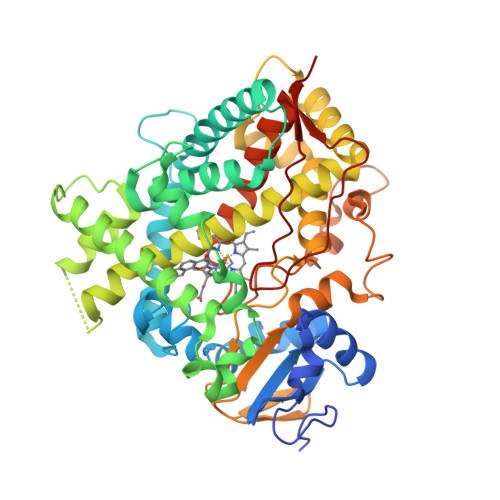
Earlier this year [1], the crystal structures of human aldosterone synthase (CYP11B2) were solved in complex with a substrate 11-deoxycorticosterone [2] and an inhibitor fadrozole [3].
Strushkevich, N., Gilep, A.A., Shen, L., Arrowsmith, C.H., Edwards, A.M., Usanov, S.A. and Park, H.-W. (2013) Structural insights into aldosterone synthase substrate specificity and targeted inhibition. Molecular Endocrinology27, 315—324.
On a number of occasions, I was chilling beer in the freezer. Then taking it out, opening the bottle and pouring it into the glass. There are four experimentally observed outcomes.
Beer is liquid both in a bottle and in a glass
Beer is liquid in a bottle but gets frozen as poured in a glass
Beer gets solid in a bottle so no way to pour it in a glass
Beer bottle explodes in the freezer
Scenario B is the most interesting one. (Scenario D is also interesting, but I won’t recommend it.) This is how the resulting beer slush looks like:
Now I saw a number of articles on the web where they explain this phenomenon with supercooling. I should say that I am not satisfied with this explanation. Why “supercooled” beer is not getting frozen in the bottle, even if I shake it, but forms slush once outside? When I put a bottle of (non-fizzy) rosé in a freezer, it either stays liquid (and remains liquid upon opening and pouring) or develops fine crystals of ice (which stay as they are upon opening both in a bottle and in a glass). On one occasion, a forgotten in a freezer bottle of rosé got frozen solid. (According to The Academic Wino, “the freezing point of table wine is –5 °C”, and my freezer goes down to –18 °C.)
To explain what happens, we don’t need to bring supercooling in. We just have to keep in mind that our drink is an aqueous solution. And that alcohol is only one of many solutes there. Of them, the most important are sugars and carbon monoxide. (Some beers, such as Guinness, contain dissolved dinitrogen as well as CO2.)
For dissolution to take place, the overall change of free energy should be negative, but the heat may be either absorbed or released. The dissolving of sugar in water is an endothermic process. The increase in temperature results in an increase in solubility. The reverse process, precipitation (often in form of crystallisation), is exothermic.
On the contrary, the dissolving of gases in water is exothermic. The increase in temperature results in an decrease in gas solubility. The reverse process, gas evolution, is endothermic. So as soon as the bottle is opened, the gas starts to escape and the temperature drops — in our example, below the freezing temperature. Sometimes, it drops so rapidly that beer freezes in the bottleneck.
And while I am on it: as this video shows, dissolving of alcohol in water is also exothermic. Not that it changes much in our beer glass experiments.
One day, idly browsing the web (as usual), I came across this:
I disagree with an unknown (to me) co-author of Antoine de Saint-Exupéry. For one thing, you don’t have to be “in organic chemistry” to recognise a reaction coordinate diagram. For another, 25 or so years ago my first reaction (that is, if I never read Le Petit Prince) would be: “Hey dude, your CD spectrum is upside down”. The fact is, I am still alive, so my life then was far from being over.
Isn’t the Web great? Nowadays I don’t have to venture to the library and sift through the J. Biol. Chem.’s and J. Mol. Biol.’s. (Even if I wanted, there is no library like that in Fuerteventura.) I can get the CD spectra online and for free in the Protein Circular Dichroism Data Bank [1]. Better still, using DichroMatch I can find spectra that are similar to my query [2]. (I just checked: it works!) Here’s how the CD spectrum of a typical α-helical protein (such as haemoglobin) looks like (a):
(a)
So... where’s a hat? Back in 1990s, our lab had a decommissioned Jobin Yvon Mark IV dichrograph, which, as I understand now, was an excellent machine. The haemoglobin spectrum would look more or less like this (b):
(b)
Neither equipment nor our samples allowed us to collect spectra below 200 nm, therefore most of the spectrum was in the negative ellipticity region. We did not really need to go below 200 nm: we were mostly monitoring ellipticity at 222 nm as a function of temperature or concentration of guanidinium chloride or other denaturing agents.
Mind you, not all proteins have this inverted hat region in their CD spectra. For example, ferredoxin (c), rubredoxin (d) or immunoglobulin G (e):
(c)
(d)
(e)
In the 21st century, protein X-ray crystallography became very much a routine technique. Once you solve the structure, there’s no mystery left. On the contrary, the CD spectra are as beautiful and enigmatic as star spectra. They still need an intelligent interpreter. They tell the story and in the same time keep the secret. I think the little prince would appreciate them.
Whitmore, L., Woollett, B., Miles, A.J., Klose, D.P., Janes, R.W. and Wallace, B.A. (2011) PCDDB: the protein circular dichroism data bank, a repository for circular dichroism spectral and metadata. Nucleic Acids Research39, D480—D486.
Klose, D.P., Wallace, B.A. and Janes, R.W. (2012) DichroMatch: a website for similarity searching of circular dichroism spectra. Nucleic Acids Research40, W547—W552.
CYP1A1 was one of the first P450 enzymes to be characterised and, as its name indicates, holds the first place in the systematic nomenclature of P450s [1]. However, it was not until last year that the first crystal structure of human CYP1A1 in complex with α-naphthoflavone has been determined at 2.6 Å resolution [2]. The structure [3] is released this week.
Naturally, you are supposed to figure this out from the first principles, or rather, from some principles taught in this course, not from Wikipedia (or “by googling”, as some put it).
(a)
(b)
The problem is, the “right” answer, PF3, is actually, factually wrong. Even though this question cost only two points (of 150), a rather animated debate followed the exam.
Those who defended the “right” but factually wrong answer (a) were proposing that what the problem was testing our thinking rather than actual knowledge, and our thinking should have been along the lines of VSEPR model. VSEPR rules correctly predict PF3 to be trigonal pyramidal and PF5 to be trigonal bipyramidal. The dipole moment of a polar molecule PF3 should make it less volatile than apolar PF5. Those who chose the “wrong” but factually correct answer (b) were arguing that polarisability of larger PF5 is higher than that of PF3 and therefore the London dispersion forces in PF5 would beat dipole-dipole interactions in PF3. The (a) party were saying that making the answer you’d get by applying principles different to the one you’d get by “googling” is a good protection against cheating. The (b) party were retorting that this is a silly way of protection, that the question asked was what has the higher boiling point, not what could be expected to have the higher boiling point, and that expecting students to come up with the factually wrong answer is not exactly pedagogical.
Truth to be told, the methods of estimating boiling or melting points of materials were simply not a part of this course. The only thing one could do was to determine whether the molecule has a non-zero dipole moment. But there is no way to figure out which effect will be stronger, the increase in dispersion forces or dipole-dipole interactions.
One would think that the physical properties of such simple compounds as binary halides of Group 15 elements (pnictogens) are studied well and long ago. Not really. I tried to compile a table of dipole moments and melting/boiling points for pnictogen tri- and pentahalides, MX3 and MX5, using various resources [1—5]. As you can see, there are still many gaps.
The dipole moment of MX3 grows larger down the group of the central atom M, e.g. μ(NF3) < μ(PF3) < μ(AsF3), and grows smaller down the group of ligand atom X, e.g. μ(SbCl3) > μ(SbBr3) > μ(SbI3).
As the sizes of both central atom and ligands go up, so do the melting and boiling points.
As dipole moments go up, so do the melting and boiling points.
Something curious happens, though, when one crosses the phosphorus—arsenic borderline. AsF3 has a dipole moment of 2.59 debye. As expected, both mp and bp of AsF3 are, respectively, higher than those of AsF5. PF3, however, has much lower moment of 1.03 D. Both mp and bp of PF3 are, respectively, lower than those of PF5. Similarly, mp of AsCl3 is higher than mp of AsCl5, whereas mp of PCl3 is lower than mp of PCl5. Similarly... but no, there are too many gaps in the table “down there”.
Which shows, by the way, that “googling” does not help if the data is not available. For the future, the course authors may consider asking a very similar question about a pair of compounds from “down there”. Thus the whole conflict between the (as yet unknown) “truth” and “expected answer” could be easily avoided.
Since the electronegativities decrease down the group for both M and L, the most polar M—L bond must be Bi—F bond and BiF3 should have the largest dipole moment. Well I couldn’t find its value. But it is known that bismuth trifluoride has ionic structure, and has the highest melting (649 °C) and boiling (900 °C) points of all binary pnictogen halides. On the other side of the spectrum, we have extremely sensitive nitrogen triiodide. A feather tickle, a loud noise and, I suppose, any attempt to measure its dipole moment will set off an explosive decomposition (see the video below):