One day, idly browsing the web (as usual), I came across this:

I disagree with an unknown (to me) co-author of Antoine de Saint-Exupéry. For one thing, you don’t have to be “in organic chemistry” to recognise a reaction coordinate diagram. For another, 25 or so years ago my first reaction (that is, if I never read Le Petit Prince) would be: “Hey dude, your CD spectrum is upside down”. The fact is, I am still alive, so my life then was far from being over.
Isn’t the Web great? Nowadays I don’t have to venture to the library and sift through the J. Biol. Chem.’s and J. Mol. Biol.’s. (Even if I wanted, there is no library like that in Fuerteventura.) I can get the CD spectra online and for free in the Protein Circular Dichroism Data Bank [1]. Better still, using DichroMatch I can find spectra that are similar to my query [2]. (I just checked: it works!) Here’s how the CD spectrum of a typical α-helical protein (such as haemoglobin) looks like (a):

(a)
So... where’s a hat? Back in 1990s, our lab had a decommissioned Jobin Yvon Mark IV dichrograph, which, as I understand now, was an excellent machine. The haemoglobin spectrum would look more or less like this (b):

(b)
Neither equipment nor our samples allowed us to collect spectra below 200 nm, therefore most of the spectrum was in the negative ellipticity region. We did not really need to go below 200 nm: we were mostly monitoring ellipticity at 222 nm as a function of temperature or concentration of guanidinium chloride or other denaturing agents.
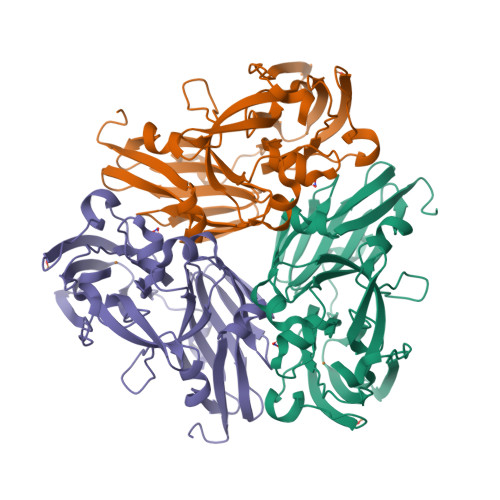
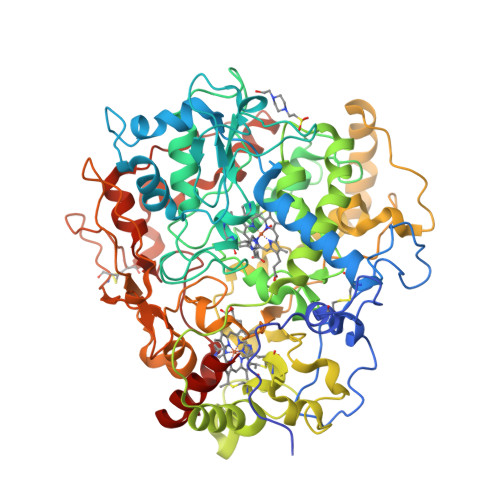
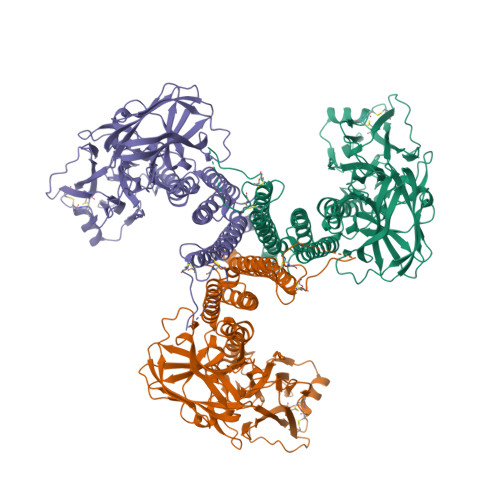
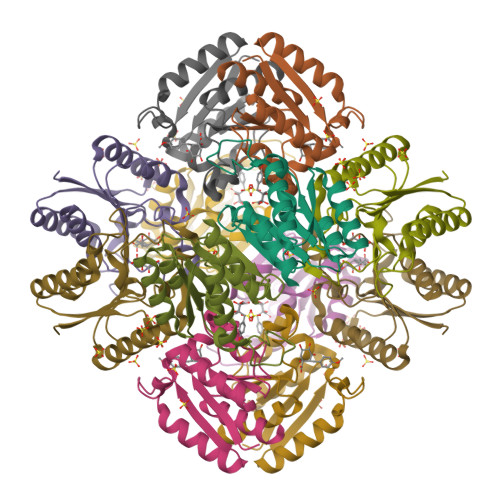
Mind you, not all proteins have this inverted hat region in their CD spectra. For example, ferredoxin (c), rubredoxin (d) or immunoglobulin G (e):

(c)

(d)

(e)
In the 21st century, protein X-ray crystallography became very much a routine technique. Once you solve the structure, there’s no mystery left. On the contrary, the CD spectra are as beautiful and enigmatic as star spectra. They still need an intelligent interpreter. They tell the story and in the same time keep the secret. I think the little prince would appreciate them.
- Whitmore, L., Woollett, B., Miles, A.J., Klose, D.P., Janes, R.W. and Wallace, B.A. (2011) PCDDB: the protein circular dichroism data bank, a repository for circular dichroism spectral and metadata. Nucleic Acids Research 39, D480—D486.
- Klose, D.P., Wallace, B.A. and Janes, R.W. (2012) DichroMatch: a website for similarity searching of circular dichroism spectra. Nucleic Acids Research 40, W547—W552.